
Written by Georgina Nicholson
Parrotfish and Cyanobacteria
Recently, the microphage feeding hypothesis proposed that microscopic photoautotrophs, particularly cyanobacteria, are the target prey for parrotfishes (Clements et al., 2017). This new theory presented an alternative view to the prevailing paradigm that parrotfish are generalist feeders on turfing algae, detritus and macroalgae.
The aim of my Masters and now my PhD is to test the microphage feeding hypothesis for parrotfish. The study of coral reef cyanobacteria has therefore been central to my research, which attempts to resolve parrotfish feeding across all potential macroscopic and microscopic prey items.
Coral Reef Cyanobacteria
Cyanobacteria (previously known as the Blue-Green Algae) can be overlooked due to their diminutive size and for this reason their presence often goes unreported on benthic surveys. Filamentous cyanobacteria are essential to healthy coral reef ecosystems due to their diazotrophy (ability to fix atmospheric nitrogen) and are integral to many coral reef turfs. Eutrophication and benthic disturbance favour these fast-growing pioneering photoautotrophs, and the well-documented degradation of coral reef ecosystems has highlighted the need to fully understand these amazing, but often neglected ancient oxygenic micro-organisms.
Cyanobacteria have important coral reef functions due to the adhesive ability of their exopolysaccharides. Cyanobacteria produce extracellular polysaccharides (Decho and Gutierrez, 2017) and which are visible in cyanobacteria as mucilaginous sheaths. Coral reef cyanobacteria create biofilms on recently exposed surfaces and bind sand grains together to stabilise and accumulate reef sand flats. Filamentous cyanobacteria are a key component of short coral reef turfs and act to trap sediment within the turf.
Lizard Island Cyanobacteria
Here I introduce two coral reef filamentous cyanobacteria that I encountered in my parrotfish prey community analysis. To identify the coral reef cyanobacteria in the epilithic prey community targeted by coral reef parrotfishes, I had to ‘bring the reef to the microscope’. Using differential interference contrast microscopy to 1000x enabled me to identify and quantify their density and investigate their epiphytic (grows on algae or other cyanobacteria), epilithic (lives on calcareous surfaces) and endolithic (lives within coral and calcareous substances) habits.
Lyngbya
Lyngbya (Oscillatoriales) is a non-heterocystous, unbranched, filamentous cyanobacteria. Lyngbya are well known for their role in both freshwater and marine blooms and identified in coral reef benthic mats (Charpy et al., 2012). This genus was the most dominant filamentous cyanobacteria I found in the prey communities targeted by five GBR parrotfish species. In my samples Lyngbya occurred unattached to other biota, although was found in association with Lobophora and mixed in with Jania. Interestingly, sticky pennate diatoms occasionally attached to their untapered trichomes.
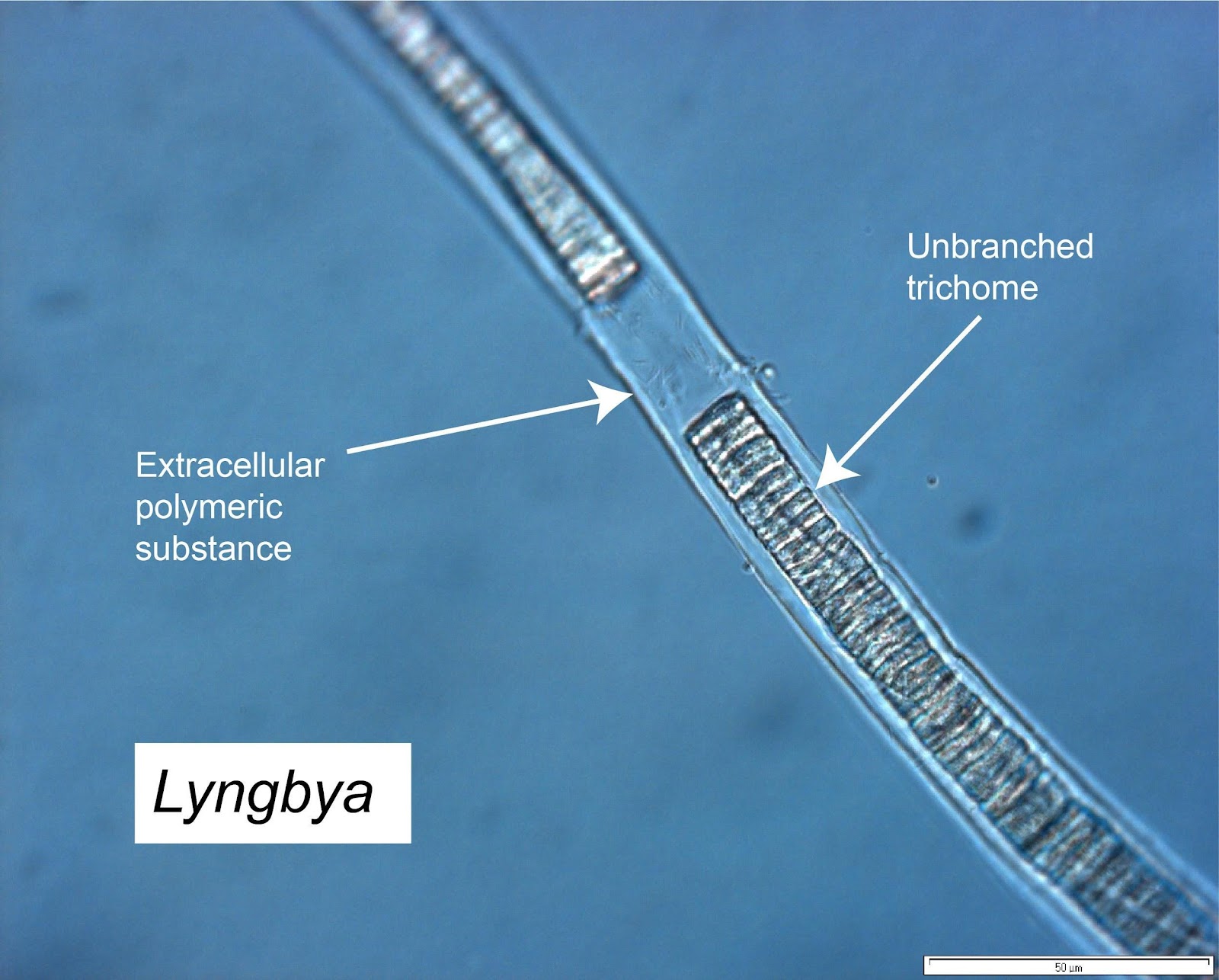
Figure 1. Lyngbya filament illustrating the mucilaginous sheath (Photo GN)
Calothrix
Calothrix (Nostocales) is an unbranched, heterocystous cyanobacteria, with a tapered trichome and visible sheath. Short filaments possess a basal heterocyst, whilst intercalary heterocysts are also present in the longer trichomes. I observed Calothrix in a variety of habits. As an epiphyte on filamentous rhodophytes attached by the basal heterocyst; also as a biofilm on crustose coralline algae, also as a tuft.
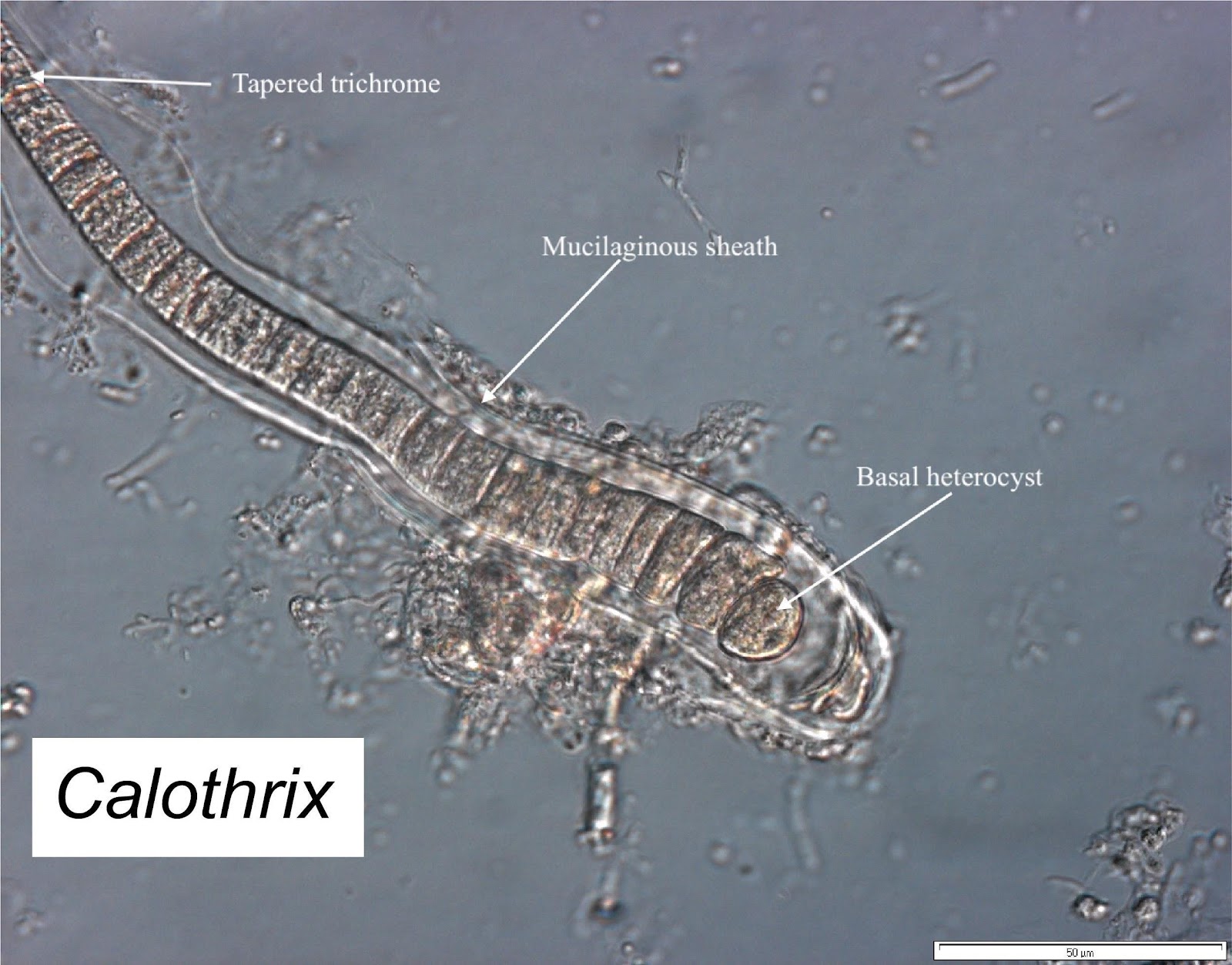
Figure 2. Calothrix, a versatile diazotrophic coral reef cyanobacteria (Photo GN).
Future Directions
Our research (paper in review) revealed the presence of filamentous cyanobacteria on all benthic samples targeted by Chlorurus and Scarus parrotfish. Additionally we found that sympatric parrotfish demonstrate partitioning of these micro-photoautotrophs at fine spatial scales. Currently I am investigating how cranial jaw morphology and jaw mechanics relates to the trophic resource partitioning we observed in the parrotfishes at Lizard Island.
Our results will be used to refine coral reef trophodynamic models, develop our understanding of parrotfish evolution, clarify the ecological roles of parrotfish, shed light on coral reef disturbance dynamics and predict how coral reefs will change under future conditions.
References
Charpy, L., Casareto, B. E., Langlade, M. J., & Suzuki, Y. (2012). Cyanobacteria in coral reef ecosystems: a review. Journal of Marine Biology. DOI: 2012.10.1155/2012/259571
Clements, K. D., German, D. P., Piché, J., Tribollet, A., & Choat, J. H. (2017). Integrating ecological roles and trophic diversification on coral reefs: multiple lines of evidence identify parrotfishes as microphages. Biological Journal of the Linnean Society, 120(4). DOI: 729-751.10.1111/bij.12914
Decho, A. W., & Gutierrez, T. (2017). Microbial extracellular polymeric substances (EPSs) in ocean systems. Frontiers in microbiology, 8, 922.DOI: 10.3389/fmicb.2017.00922.
