
Written by Charlotte Page
Summary: Driving patterns of coral bleaching (loss of their algae symbionts) over reefs are a suite of biophysical interactions where the physical environment shapes organism response through an interplay with intrinsic biological functioning. In this article I will explore different aspects of a reefs physical environment which can in turn affect the biology of corals and therefore their responses to thermal stress. These are ideas that we discuss in depth in our recent review article published in the journal BioEssays.

The physical oceanographic environment shapes the distributions and livelihoods of many marine species. The movement of water around the globe on large and small scales, sunlight penetration, water temperature, and ocean chemistry – all affect the kind of life an organism might live and where.
Corals as benthic marine animals spend a majority of their lives cemented in one place, and therefore seem to really live at the whim of their physical surroundings. And when we consider how harsh this physical environment seems to be – often sunny, warm and low in food sources, we can begin to see why corals are sensitive to environmental change, where any variation could be detrimental and tip them over the edge.

When we think about mechanisms that can cause the coral-algal relationship to break down, it always exists as a response to the manifestation of some sort of physical stress outside the normal range a coral may function happily within – the obvious offender here being climate change induced ocean warming. The accumulation of anomalously high summer water temperatures relative to their historical norm disrupts the coral-algal symbiosis, causing coral bleaching which most often leads to coral death. In this case the physical environment is interacting with the biological functioning of the coral ‘meta-organism’ (the coral host and all associated symbiotic microbiota) in a wholly negative way. But like any good story, where there lies a monster, there also lies a hero.
The thermal environment a coral experiences is often the key to determining the ‘biological’ outcome in response to stress (i.e. severity of bleaching or other stress response). At the reef scale during broad summer temperature anomalies we see variation in thermal environment across different reef habitats causing different outcomes. For example in 2015 the Dongsha Atoll saw up to 70% mortality in reef flat areas compared to reef slope and channel environments. This was due to a 6 °C amplification (see Figure 1 below) of local temperatures on the reef flat caused by a multitude of physical factors. Equally the thermal environment can itself induce physiological changes in the coral metaorganism through acclamatory mechanisms, for instance corals exposed to variability in temperature prior to heating events can develop an increased tolerance to heat stress.
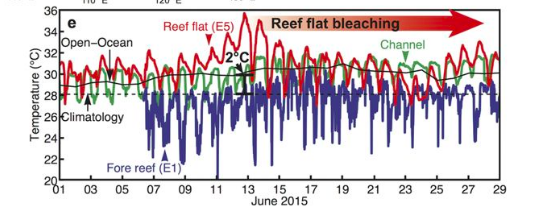
Consequently it becomes clear that an ability to predict and characterise the thermal environment over reefs and factors which can cause variability therein is crucial. The thermal environment over reefs can be accurately estimated through the use of heat budgets. Temperature variations experienced at a location depend on the net rate of heat transfer into and out of the water. Heat budgets include a number of parameters to estimate the net effects of these fluxes, including exchanges from the sun and the surface of the ocean, in addition to the transfer of heat within the ocean itself.
Currents distribute heat over reefs through both advection (the horizontal movement of water) or convection (up and down movement of water). Currents can therefore directly effect the thermal environment over reefs, meaning that they inherently play a role in determination of bleaching risk. These impacts scale widly, from the effects of local mixing reducing stratification of the water column, to that of large scale upwelling patterns and oceanographic currents.
As we zoom in from whole reefs, to that of individual corals, we find that water flow also has impacts on the biological functioning of reefs through modulation of physiology. Corals regulate the gases and metabolites they take up and dispel through passive diffusion. This process occurs over what is called ‘boundary’ layers. Boundary layers emerge through the interaction of the coral’s surface with the viscous forces in fluid. Gases, as well as other important metabolites, move across this layer and so regulate the corals’ cellular physiology ( described in Figure 2). The thickness of this layer directly effects the time diffusion takes, and therefore the rates of many physiological processes.
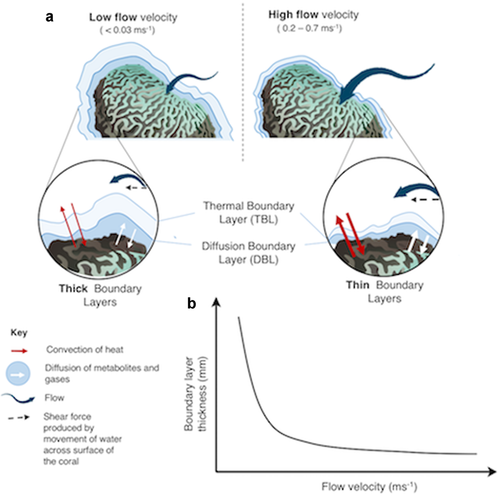
Under higher flow conditions boundary layers become thinner, causing increased rates of transfer of these gases and metabolites from the coral tissue to the water columns and vice versa. Higher flow conditions have therefore been experimentally related to higher rates of a number of physiological parameters, including respiration, photosynthesis and calcification. At this level higher flow conditions have also been hypothesised as directly controlling rates of photo-oxidative damage during temperature stress.
There is also evidence of flow being a beneficial physical factor ecologically too. Flow conditions have been recorded as having positive effects ecologically during bleaching events, where higher flow condition corals seem to be ‘healthier’ during summer time heating events, leading to less prevalence of bleaching.

This is all very well but ultimately what we are working towards is understanding how physical factors like flow will contribute towards future reef resilience under a changing climate. This knowledge can also help in finding practical solutions to the coral bleaching crisis – if we know how corals can respond to temporal and spatial variability in the physical environment, then oceanographic modelling can be integrated with our knowledge of cellular physiology. This link underlies the degree heating week approach used commonly within the field to understand bleaching risk. Without a clearer understanding of the rules that define biological responses to climate change, science and management will forever be playing catch up in a world where we need to start strategically planning more than we are reactively responding.
References:
Comeau S, Edmunds P, Lantz C, and Carpenter R. 2014. Water flow modulates the response of coral reef communities to ocean acidification. Scientific reports 4:6681.
DeCarlo TM, Cohen AL, Wong GT, Davis KA, Lohmann P, and Soong K. 2017. Mass coral mortality under local amplification of 2 C ocean warming. Scientific reports 7:44586.
DeCarlo TM, and Harrison HB. 2019. An enigmatic decoupling between heat stress and coral bleaching on the Great Barrier Reef. PeerJ 7.
Nakamura T, and van Woesik R. 2001. Water-flow rates and passive diffusion partially explain differential survival of corals during the 1998 bleaching event. Marine Ecology Progress Series 212:301-304.
Nakamura T, Van Woesik R, and Yamasaki H. 2005. Photoinhibition of photosynthesis is reduced by water flow in the reef-building coral Acropora digitifera. Marine Ecology Progress Series 301:109-118.
Osinga R, Derksen-Hooijberg M, Wijgerde T, and Verreth JA. 2017. Interactive effects of oxygen, carbon dioxide and flow on photosynthesis and respiration in the scleractinian coral Galaxea fascicularis. Journal of Experimental Biology:140509.
Page CE, Leggat W, Heron SF, Choukroun SM, Lloyd J, and Ainsworth TD. 2019. Seeking Resistance in Coral Reef Ecosystems: The Interplay of Biophysical Factors and Bleaching Resistance under a Changing Climate. BioEssays 41:1970071. 10.1002/bies.201970071
Schoepf V, Stat M, Falter JL, and McCulloch MT. 2015. Limits to the thermal tolerance of corals adapted to a highly fluctuating, naturally extreme temperature environment. Scientific reports 5:17639. 10.1038/srep17639
https://www.nature.com/articles/srep17639 – supplementary-information
Skirving W, Heron M, and Heron S. 2006. The Hydrodynamics of a Bleaching Event: Implications for Management and Monitoring. Coral Reefs and Climate Change: Science and Management: American Geophysical Union, 145-161.
